
La especie humana posee 46 cromosomas dispuestos en 23 pares, de esos 23 pares 22 son somáticos o autosomas (heredan caracteres no sexuales) y uno es una pareja de cromosomas sexuales (llamados también heterocromosomas o gonosomas), identificados como XX en las mujeres y como XY en los hombres.
Esta pareja de cromosomas sexuales no solo llevan los genes que determinan el sexo, sino que también llevan otros que influyen sobre ciertos caracteres hereditarios no relacionados con el sexo.
Hay caracteres que sin ser caracteres sexuales primarios (órganos genitales, gónadas) o secundarios (barba del hombre, pechos de las mujeres), solo aparecen en uno de los dos sexos, o si aparecen en los dos, en uno de ellos son mucho más frecuentes.
A estos caracteres se les denomina caracteres ligados al sexo.
Variantes en la herencia ligada al sexo
Vimos que los cromosomas sexuales constituyen un par de homólogos (XX en la mujer y XY en el hombre); sin embargo, en el par XY un segmento de cada cromosoma presenta genes particulares y exclusivos (segmento heterólogo, llamado también diferencial o no homólogo), la porción restante de los cromosomas del par XY corresponde al sector homólogo, como se grafica en el esquema siguiente:
 |
.
Los varones sólo llevan un representante de cada gen ubicado en el sector heterólogo del X (en tanto poseen un X) y las mujeres portan dichos genes por pares (en tanto poseen dos X). Por consiguiente, la transmisión y expresión de estos genes dependen del sexo de los individuos.
Entonces, la herencia ligada al sexo se refiere a la transmisión y expresión, en los diferentes sexos, de los genes que se encuentran en el sector no homólogo (heterólogo) del cromosoma X heredado del padre.
También podemos decir que la herencia ligada al sexo no es más que la expresión en la descendencia de los genes ubicados en aquellas regiones del cromosoma X que no tienen su correspondencia en el cromosoma Y.
En el sexo femenino, la presencia de dos cromosomas X hace que los genes contenidos en estos se comporten como si se encontraran en autosomas, con normalidad.
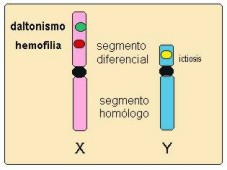 |
| Daltonismo y hemofilia, dos anomalías recesivas ubicadas en el segmento diferencial del cromosoma X. |
Así, pues, un carácter determinado por un gen del cromosoma X aparecerá si la mujer tiene un alelo dominante en cada uno de estos cromosomas, o si tiene dos alelos recesivos, uno en cada uno de ellos (homcigota en ambos casos).
Si, en cambio, la mujer es heterocigota para ese carácter, igual se manifestará el alelo dominante. Es decir, se trata de un modelo de herencia clásico y normal, comparable a los mencionados en las leyes de Mendel.
El caso del hombre es radicalmente distinto. Si los genes se encuentran en la zona del cromosoma X que tiene su parte correspondiente (homóloga) en el Y, actúan como en el caso anterior.
De hecho, se han encontrado algunos alelos en los sectores homólogos de los cromosomas XY (llamados genes pseudoautosómicos) y dadas las características de estas regiones especiales de los cromosomas sexuales, los trastornos producidos a causa de estos genes se heredarán de acuerdo a las leyes genéticas que rigen la herencia autosómica (o sea, funcionan igual que los autosómicos).
Pero es mucho más frecuente que los genes estén en una parte del cromosoma X que no tenga correspondencia en el Y. Si esto sucede, los alelos se manifestarán siempre, ya sean dominantes o recesivos.
Por tanto, debemos recordar que los genes ligados a los cromosomas sexuales pueden ser tanto recesivos como dominantes.
HERENCIA RECESIVA LIGADA AL SEXO
En los hombres, los cromosomas X e Y se pueden aparear durante la meiosis (en la Profase I) por sus fragmentos homólogos y por tanto se pueden producir entrecruzamientos en esta zona.
Herencia ligada al cromosoma Y o herencia holándrica
Todos los genes que se encuentran en el segmento diferencial o no homólogo del cromosoma Y son heredados únicamente por los hijos varones y se manifestarán en todos los hombres que los lleven y sólo en los hombres, independientemente de que sean dominantes o recesivos.
La transmisión de los genes situados en el segmento no homólogo del cromosoma Y es la herencia holándrica..
Holándrico:
(Del griego olos, todo, y alter, hombre). Dícese de la transmisión hereditaria de una tara o de una enfermedad que se realiza de un padre a todos sus hijos, quedando indemnes las hijas. Está ligada a genes situados en el segmento no homólogo del cromosoma sexual Y constituyéndose en una variedad de herencia ligada al sexo.
 |
| Thomas Hunt Morgan (25 de septiembre 1866 - 4 de diciembre 1945), genetista estadounidense, quien concluyó que algunos caracteres se heredan ligados al sexo. |
Herencia ligada al cromosoma X.
La herencia ligada al cromosoma X quiere decir que el gen que causa el rasgo o el trastorno se localiza en el cromosoma X .
Cabe recordar que las mujeres poseen dos cromosomas X mientras que los hombres poseen un cromosoma X y un cromosoma Y. Los genes del cromosoma X pueden ser recesivos o dominantes, y su expresión en las mujeres y en los hombres no es la misma debido a que los genes del cromosoma Y no van apareados exactamente con los genes del X.
Los genes recesivos ligados al cromosoma X se expresan en las mujeres únicamente si existen dos copias del gen (una en cada cromosoma X). Sin embargo, en los varones sólo debe haber una copia de un gen recesivo ligado al cromosoma X para que el rasgo o el trastorno se exprese.
Por ejemplo, una mujer puede ser portadora de un gen recesivo en uno de sus cromosomas X sin saberlo y transmitírselo a su hijo, que expresará el rasgo o el trastorno.
Entre los ejemplos de trastosnos recesivos ligados al cromosoma X se destacan los casos del daltonismo y la hemofilia, enfermedades provocadas por un gen recesivo situado precisamente en el segmento diferencial del cromosoma X.
Recalcamos que, debido a su ubicación, para que una mujer padezca la enfermedad debe ser homocigota recesiva (tener el gen recesivo en ambos cromosomas X), mientras que en los hombres basta con que el gen recesivo se encuentre en el único cromosoma X que tienen.
Daltonismo
Esta enfermedad, determinada por un gen recesivo del cromosoma X, es una anomalía que consiste en la incapacidad de distinguir los colores rojo y verde. Se suele llamar también ceguera para los colores, y hay muchos tipos.
La enfermedad fue descrita por una persona afectada, el químico inglés John Dalton, en 1794. El nombre de esta alteración hace referencia, precisamente, a este científico.
Como ya dijimos, el gen responsable de la enfermedad es recesivo y su presencia origina el daltonismo en el hombre, mientras que la mujer que lo posee es portadora y no lo manifiesta. Para que una mujer sea daltónica es necesario que tenga genes del daltonismo en los dos cromosomas X (homocigota) , lo cual es bastante poco frecuente.
La agudeza visual (la capacidad de ver) del daltónico es normal. No existen grandes complicaciones; sin embargo, los individuos afectados pueden no ser considerados para ciertos empleos relacionados con el transporte o las Fuerzas Armadas en donde es necesario el reconocimiento de colores.
Este trastorno es 16 veces más frecuente en los hombres que en las mujeres, debido a que el gen se localiza en el cromosoma X (que es uno solo en el hombre y son dos en las mujeres; por tanto; la mujer tiene mayores posibilidades de tener el gen dominante para la visión normal en uno de los cromosomas X).
Recordemos que dominante es el alelo que se expresa y se designa con mayúscula.
Recordemos, además, que el alelo recesivo sólo se expresa cuando no está presente el dominante y se designa con minúscula.
Si caracterizamos con una letra N mayúscula (normal) la condición del cromosoma que lleva el el gen dominante que caracteriza la visión normal (sin daltonismo en nuestro caso), y con una d minúscula la tenencia del cromosoma con el el gen recesivo para el daltonismo, las posibles combinaciones que se pueden dar son:
(Recordemos que el cromosoma Y es más corto que el X y por ende es vacío en la parte donde se halla el gen que determina la visión normal).
1. Madre normal (XNXN) y padre normal (XNY):
| XN | XN | |
| XN | XNXN | XNXN |
| Y | XNY | XNY |
Ninguno de sus hijos (hombres y mujeres) será daltónico ni portador.
2. Madre normal (XNXN) y padre daltónico (XdY):
| XN | XN | |
| Xd | XdXN | XdXN |
| Y | XNY | XNY |
Todas las hijas portadoras (100 por ciento) y todos los hijos hijos normales (100 por ciento).
La combinación descrita se grafica en la imagen siguiente, donde el gen recesivo del daltonismo se identifica con la equis en rojo (X):
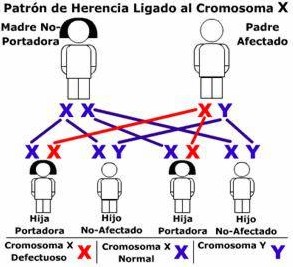 |
| Solo hijas portadoras, hijos no afectados. |
3. Madre portadora y padre normal:
| Xd | XN | |
| XN | XNXd | XNXN |
| Y | XdY | XNY |
El 50 por ciento de sus hijas seran portadoras y el 50 por ciento de sus hijos seran daltónicos.
La combinación descrita se grafica en la imagen siguiente, donde el gen recesivo del daltonismo se identifica con la equis en rojo (X):
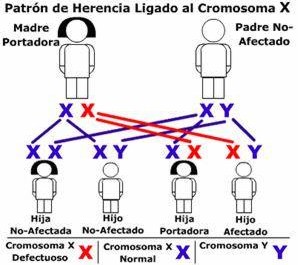 |
| Hijas e hijos, unos sanos, otros portadores. |
4. Madre portadora y padre daltónico:
| Xd | XN | |
| Xd | XdXd | XdXN |
| Y | XdY | XNY |
El 50 por ciento de hijas portadoras, 50 por ciento hijas daltónicas, 50 por ciento hijos daltónicos, 50 por ciento hijos normales.
5. Madre daltónica y padre normal:
| Xd | Xd | |
| XN | XNXd | XNXd |
| Y | XdY | XdY |
Todas las hijas portadoras y todos los hijos daltónicos.
6. Madre daltónica y padre daltónico:
| Xd | Xd | |
| Xd | XdXd | XdXd |
| Y | XdY | XdY |
Todos los hijos (hombres y mujeres) daltónicos.
Estadísticamente, lo más habitual es la madre portadora con un padre normal.
Analicemos un caso:
¿Qué tipo de visión tendrá la descendencia de una mujer de visión normal cuyo padre era daltónico (por lo tanto es portadora) y un hombre daltónico cuyo padre era también daltónico?
¿Cuáles serám sus genotipos y fenotipos?
Solución:
Sabemos que el daltonismo depende de un gen recesivo localizado en el segmento diferencial del cromosoma X.
Para esta enfermedad las mujeres pueden ser homocigóticas (XNXN o XdXd) o heterocigóticas (XNXd).
Debidos a que el alelo que determina el daltonismo es recesivo (d), las mujeres heterocigóticas son portadoras sin padecerla y las homocigóticas son daltónicas. Por el contrario, los hombres sólo pueden ser homocigóticos para la enfermedad (XNY) o (XdY), por lo que, si portan el alelo recesivo causante del daltonismo, siempre la padecen.
Partiendo de esta explicación el planteamiento del problema es el siguiente:
Identifiquemos los cromosomas como
N: normal
d: daltonismo
Partiendo que la mujer es de fenotipo normal (no manifiesta la enfermedad) pero de padre daltónico el único genotipo posible es: XNXd (portadora)
En el caso del hombre, siendo éste daltónico y de padre daltónico su genotipo es: XdY
Entonces, sus combinacines son:
| XN | Xd | |
| Xd | XNXd | XdXd |
| Y | XNY | XdY |
Los genotipos y fenotipos de la descendencia son:
De las mujeres, existe el 50 por ciento de probabilidades que sean de fenotipo normal (genotipo XNXd) y otro 50 por ciento que sean de fenotipo daltónicas (genotipo XdXd).
De los hombre, existe el 50 por ciento de posibilidades de que sean de fenotipo normal (genotipo XNY) y otro 50 por ciento de que sean daltónicos (XdY).
Ver: PSU: Biología; Pregunta 10_2006
Hemofilia A
La hemofilia A es un trastorno en el cual la sangre no coagula adecuadamente debido a una insuficiencia del factor de coagulación llamado Factor VIII. El resultado es un sangrado abundante anormal que no se detiene, aun en el caso de una cortadura pequeña.
A las personas con hemofilia A les aparecen moretones con facilidad y pueden tener hemorragias internas dentro de las articulaciones y los músculos.
La hemofilia A ocurre en uno de cada 10.000 varones recién nacidos.
Existe un tratamiento mediante la infusión del Factor VIII (transfusión de sangre).
Las mujeres portadoras del gen pueden mostrar signos leves de la insuficiencia del Factor VIII como los moretones que aparecen con facilidad o las hemorragias que tardan más de lo normal en detenerse luego de una cortadura. Sin embargo, no todas las mujeres portadoras presentan estos síntomas.
Se cree que un tercio de todos los casos son nuevas mutaciones en la familia (no heredadas de la madre).
Herencia dominante ligada al sexo
La herencia dominante ligada al sexo tiene lugar cuando un gen anormal de uno de los padres es capaz de causar la enfermedad, aunque pueda haber un gen compatible proveniente del otro padre que sea normal. El gen anormal domina el resultado del par de genes.
Aunque la gran mayoría de las enfermedades que en la especie humana son transmitidas por genes ligados al sexo se deben, como ya vimos, a un gen recesivo, también hay ejemplos en los cuales el gen para alguna de las enfermedades es dominante, aunque los genes dominantes ligados al cromosoma X son muy poco frecuentes.
Se trata de un tipo de herencia que se caracteriza por que los varones afectados transmiten el carácter a todas sus hijas y a ninguno de sus hijos (ya que estos no reciben el cromosoma X sino que el Y, que no lleva el gen. Las mujeres afectadas, en tanto, lo transmiten a la mitad de sus hijos y a la mitad de sus hijas.
A modo de ejemplo, si hay cuatro hijos (dos varones y dos mujeres) y la madre está afectada (un X anormal y tiene la enfermedad), pero el padre no, la expectativa estadística es de:
Dos hijos (una mujer y un varón) con la enfermedad
Dos hijos (una mujer y un varón) sin la enfermedad
| XE | Xs | |
| Xs | XEXs | XsXs |
| Y | XEY | XsY |
Para mayor comprensión identificamos al gen o elelo dominante con una E mayúscula (Enfermedad) y sano o recesivo con una s minúscula (sano).
Ahora, si hay cuatro hijos (dos mujeres y dos varones) y el padre está afectado (un X anormal y tiene la enfermedad), pero la madre no, la expectativa estadística es de:
Dos niñas con la enfermedad
Dos niños sin la enfermedad
| Xs | Xs | |
| XE | XEXs | XEXs |
| Y | XsY | XsY |
Un ejemplo de este tipo de herencia es la hipofosfatemia (raquitismo que no cede con la administración de vitamina D).




















.png)